Reseptör bağlanma bölgesi mimarisinin koronavirüs S, koronavirüs HE ve influenza virüsü HEF glikoproteinleri boyunca korunması. Koronavirüsler insanlarda solunum yolu enfeksiyonlarına ve dünya çapında ölümcül pnömoni salgınlarına neden olur. Enfeksiyonlar, konakçı reseptörlere bağlanan ve viral ve hücresel zarları kaynaştıran transmembran başak (S) glikoprotein tarafından başlatılır. Oligosakkarit reseptörlerine koronavirüs bağlanmasının moleküler temelini anlamak için, koronavirüs OC43 S glikoprotein trimerin kriyo-EM yapılarını izole ve 9- O ile kompleks halinde belirledikasetillenmiş siyalik asit. Ligandın hızlı kinetiklerle yüzeye maruz kalan bir oluğa bağlandığını ve belirlenen bölgedeki etkileşimlerin konakçı hücrelere S aracılı viral giriş için gerekli olduğunu, ancak serbest monosakkaridin füzojenik konformasyonel değişiklikleri tetiklemediğini gösteriyoruz.
****
****
Tüm koronavirüs korunmuş olan reseptör etkileşen Alanı 9- angaje glikoproteinleri S O koronavirüs hemaglutinin esteraz ve grip virüsü C / D hemaglutinin-esteraz füzyon glikoproteinlerin ligand bağlayıcı cepler benzer bir yapıyla, asetil-sialogycans. Sonuçlarımız, bu virüslerin sialoglikanları hedef hücrelerin yüzeyinde tutmak için benzer stratejiler geliştirdiğini göstermektedir.
Koronavirüsler, Nidovirales düzeninde büyük, pozitif anlamda zarflanmış RNA virüsleridir ve dört türe ayrılır: α, β, γ ve δ. İki β-coronavirüsler 21 yılı başından bu yana insanlarda ölümcül pnömoni salgına neden olmuş st yüzyıl. Şiddetli akut solunum sendromu koronavirüsü (SARS-CoV) 2002 yılında ortaya çıkmıştır ve 2003 yılında tutulmadan önce ölüm oranı% 10 olan beş kıtaya yayılan bir salgından sorumlu olmuştur (2004’te bildirilen ek vakalarla). Ortadoğu solunum sendromu koronavirüsü (MERS-CoV) 2012 yılında Arap Yarımadası’nda ortaya çıktı ve ölüm oranı% 35 olan insanlarda tekrarlayan salgınlara neden oldu. SARS-CoV ve MERS-CoV, yarasalar / palmiye misk kedileri 1 ve dromedary develer kullanarak tür bariyerini geçen zoonotik virüslerdir2 . Zoonotik kökenli diğer dört koronavirüs, insan popülasyonunda endemiktir ve hafif solunum yolu enfeksiyonlarının% 30’una kadarını oluşturur ve küçük çocuklar, yaşlılar ve bağışıklığı baskılanmış kişilerde ciddi komplikasyonlara veya ölümlere neden olur 3 , 4 . Bu virüsler HCoV-NL63 ve HCoV-229E (a-koronavirüsler) ve HCoV-OC43 ve HCoV-HKU1’dir (β-koronavirüsler). Şu anda, herhangi bir insan koronavirüsü ile savaşmak için spesifik bir antiviral tedavi veya aşı mevcut değildir.
***
***
Ayrıca, yarasalar 5 , 6 , 7’de bulunan büyük rezervuar göz önüne alındığında, koronavirüslerin gelecekteki çapraz tür iletim olayları muhtemel görünmektedir.. Dolayısıyla koronavirüslerin incelenmesi, türler arası iletim ve insanlara adaptasyon ile ilgili prensiplerin anlaşılmasına ve gelecekteki zoonotik salgınların hazırlanmasına yardımcı olacaktır.
Koronavirüsler, giriş için viral ve hücresel zarların konakçı bağlanmasını ve füzyonunu teşvik etmek için başak (S) glikoproteinin homotrimerlerini kullanır. S, viral yüzeyde bulunan ana antijendir ve enfeksiyon sırasında antikorları nötralize etmenin hedefidir. Sonuç olarak, aşı tasarımının bir odağıdır. S yaklaşık 1.300 amino asit 8 tek bir polipeptit zincir öncüsü olarak sentezlenen bir sınıf I viral füzyon proteinidir . Birçok koronavirüslerin için, S S adı, iki alt birimi oluşturmak için konakçı proteazları tarafından işlenir 1 S 2 , kovalent olmayan füzyon öncesi konformasyonunda bağlı olarak kaldığı. N-terminali S 1alt birim, farklı koronavirüslerde reseptör bağlanma domenleri olarak işlev gören A veya B domeniyle A, B, C ve D olarak adlandırılan dört β açısından zengin alan içerir. Transmembran C-terminali S 2 alt-birim metastabil yaylı füzyon makine olup 9 . Giriş sırasında, S 2 ayrıca proteolitik S ayrıştırı- 2’nin füzyon peptidinin hemen üst akışında, ‘sitesi 10 . Bu ikinci bölünme adımı tüm koronavirüsler için meydana gelir ve geri dönüşümsüz konformasyonel değişiklikler yoluyla gerçekleşen membran füzyonu için proteini aktive ettiğine inanılır 11 , 12 , 13 , 14. Son yıllarda, kriyo-EM çalışması, füzyon reaksiyonunun başlangıç ve bitiş noktalarının anlık görüntülerini sağlayan, füzyon öncesi ve füzyon sonrası durumlarda koronavirüs S glikoprotein ektodomain yapılarının belirlenmesine yol açmıştır 9 , 13 , 15 , 16 , 17 , 18 , 19 , 20 , 21 , 22 , 23 , 24 . İnsan nötralize edici antikorlar ile kompleks içinde SARS-CoV ve MERS-CoV S glikoproteinlerin Cryo-EM yapıları da füzyon aktivasyon 25 mekanizması hakkında bilgi .
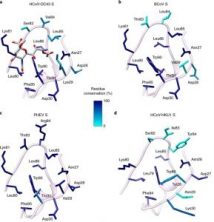
HCoV-OC43 ilk kez 1967 yılında Birleşik Krallık’taki Salisbury’deki Soğuk Algınlığı Ünitesindeki gönüllülerden izole edildi. Genom dizilerinin moleküler saat analizi, HCoV-OC43’ün bir sığır koronavirüsünün (BCoV) zoonotik iletim olayından kaynaklandığını ve 1890’lar ve 1950’ler 26 , 27 arasında en son ortak atalarını tarihlendirdiğini gösterdi . HCoV-OC43 HCoV-HKU1, BCoV ve domuz hemaglutinleyici ensefalomiyelit virüsü (PHEV) kullanımı 9- O -asetil-sialik asit (9- O -AC-Sia) terminal oligosakaritler de glikoproteinlerin ve gangliosidler dekorasyon bağlı olan bir reseptörü olarak konak hücre yüzeyi 28 , 29 . Bu virüslerin S glikoproteini 9- O’ya aracılık eder-AC-Sia bağlanma, hemaglutinin-esteraz ise (HE) protein reseptör imha edici enzim aracılığıyla sialate- olarak hareket O enfekte olmuş hücrelerden virüs soyu salımını kolaylaştırır ve non-permisif konakçıya bağlantı kaçmasına asetil-esteraz etkinliği, hücreler veya tuzakları 30 , 31 , 32 , 33 . Bu özellikler, influenza C ve D virüslerinin 28 , 34 , 35 , 36 hemaglutinin-füzyon-esteraz (HEF) glikoproteinleri ile paylaşılmaktadır .
Sialik asitler, glikokonjugatların her yerde bulunan terminal tortularıdır ve çekirdek N-asetil nöraminik asit molekülünün modifikasyonları ve glikozidik bağlantılardaki farklılıklar 37 , 38 , 39’un bir sonucu olarak geniş bir çeşitlilikte ortaya çıkar . Önceki biyokimyasal çalışma, koronavirüs S glikoproteinlerinin A alanının, 9- O -Ac-Sia 28 , 40 , 41 veya MERS-CoV ile etkileşime giren HCoV-OC43 ve BCoV gibi oligosakkarit reseptörlerine bağlanmaya aracılık ettiğini tespit etti. a2,3-bağlı (ve daha az ölçüde a2,6-bağlı) sialik asitler; sülfatlanmış sialil-Lewis X, tercih edilen bağlayıcı 42’dir.. Koronavirüs S alan A’nın galektin benzeri katına ve mutasyon analizlerine dayanarak, anahtar sakarit bağlayıcı kalıntıların BCoV sandwich-sandviçin viral membran distal tarafına yerleştirildiği ileri sürülmüştür. Bununla birlikte, son çalışmalarımız, HCoV-OC43, HCoV-HKU1, BCoV ve PHEV’in 9 – O -Ac-Sia bağlanma bölgesinin, bu virüsler arasında korunduğunu ve A 43 alanının farklı bir yerinde bulunduğunu gösterdi . Bulguları mutajenez ve BCoV enfektivite tayinleri kullanarak doğrulasak da, siliko modellemenin dışında sakkaridlerle koronavirüs etkileşimi mekanizması hakkında hiçbir yapısal bilgi mevcut değildir 43. Bu bilgi boşluğu, bu reseptörlerin viral enfeksiyon veya zoonozdaki rolleri hakkındaki anlayışımızı sınırlar ve inhibitörlerin rasyonel tasarımını engeller.
Koronavirüslerin konakçı hücrelerin yüzeyindeki siyalik asitlere bağlanmasını anlamak için, stabilize edilmiş bir HCoV-OC43 S glikoprotein trimerinin kriyo-EM yapılarını izole ve 5-N-asetil, 9 – O -asetil-nöraminik asit ile kompleks halinde belirledik 2,9-Å ve 2,8-Å çözünürlükte a-metil glikozit (9 – O -Ac-Me-Sia). Ligandın, A alanının yüzeyinde yer alan HCoV-OC43 S üzerindeki bir olukta hızlı birleşme / ayrılma kinetikleri ile bağlandığını gösteririz. 9- O -Ac-Sia bağlanması, konakçı hücrelere HCoV-OC43 S aracılı giriş için gereklidir. Sonuçlarımız ayrıca 9- O serbest bağlanmasının-Ac-Me-Sia ve / veya asidik pH, S’nin füzojenik konformasyonel değişikliklerini indüklemedi, bu da sialoglikanlarla çok değerli etkileşimlerin ve / veya varsayılan bir proteinli reseptöre 44 daha fazla bağlanmanın zar füzyonunu teşvik etmek için gerekli olduğunu düşündürmektedir . Reseptör ile etkileşen bölge, 9 – O -Ac-sialoglikanlara bağlandığı bilinen tüm koronavirüs S glikoproteinlerinde korunur ve böylece koronavirüs HE’lerinin ve influenza virüsü C / D HEF glikoproteinlerin ligand bağlayıcı cepleriyle mimari benzerliği paylaşır, böylece ortak yapısal ilkeleri vurgular tanıma 45 , 46 .
Sonuç olarak Apo-HCoV-OC43 S glikoproteininin yapısı
Biz S olduğu bir apo-HCoV-OC43 S, dış bölge trimer, mutant bir 2,9-Â çözünürlük cryo-EM yeniden tespit 1 / S 2 furin klevaj biyogenez sırasında proteolitik işlenmesini önlemek için kaldırılmıştır. HCoV-OC43 S, 150-Å yüksekliğinde ve 130-Å genişliğinde kompakt bir trimer olarak katlanır (Şekil 1a , Ek Şekil 1a, b ve Tablo 1 ). S 1 alt-birimi benzer diğer β-koronavirüs S yapılara dört alan (A, B, C ve D), 3D düzenlenmesinden elde edilen V-şekilli bir yapıya sahiptir 9 , 17 , 18 , 19 , 20 , 21 , 22 ,23 , 24 , 25 . S 2 çok S den muhafaza edilir alt-birimi, 1 koronavirüsler arasında alt birimi, C-terminal ucunda, ekli bir bağlayıcı alanı ile trimerik birimi uzatılmış bir çok sarmal olarak kıvrımlar 9 , 16 (Şek. 1a ). Mevcut koronavirüs yapıları glikoprotein S arasında, HCoV-OC43 S fare hepatit virüsü (MHV) S en çok benzeyen 9 S (rms sapması (RMSD) 979 hizalanmış Ca pozisyonlara 4,7 A) ve HCoV-HKU1’in için 18(rmsd 4.5 Å, 949 hizalı Cα pozisyonunda), sırasıyla% 62 ve% 68 sekans özdeşliğini paylaşır. Kriyo-EM rekonstrüksiyonu, her bir protomerin yüzeyinden uzanan 14 N-bağlı glikanları çözer. HCoV-OC43 S oligosakkarit yoğunluğu, SARS-CoV S ve MERS-CoV S ile karşılaştırılabilir, her üç virüs de β-cinsine aittir, ancak domuz delta coronavirus S (gen-cinsi) glikan yoğunluğundan daha düşüktür. veya HCoV-NL63 S (a-cinsi) glikoproteinleri
